ABSTRACT/INTRO:
The genus Ribes is known to be particularly diverse in western North America, with the region containing a large number of species representing many of the major lineages within the genus. However, this striking diversity is also challenging to taxonomists, as many of the species are closely related to each other and possess strong morphological similarities, with the differences between the taxa being subtle and hard to interpret.
The group of species that this study is about is rather exemplary of this issue. The group in question is one that I've informally dubbed the "desert gooseberries", and the name is informal because it has not been recognized as a singular, cohesive group in any of the literature that I've read, despite the fact that morphological and ecological factors indicate a shared common ancestor to unite these obviously similar species. The currently described species in this group include:
-
R. microphyllum, a pink-flowered species native to Mexico and far north Guatemala;
-
R. madrense, a (seemingly questionable) pink-flowered species native to Durango in Mexico;
-
R. leptanthum, a white-flowered species native to the more southern and eastern extents of the Rockies and the Colorado Plateau;
-
R. quercetorum, a yellow-flowered species native to coastal southern California and some of Baja;
-
R. lasianthum, a yellow-flowered species native to alpine communities of the high Sierras;
-
R. velutinum, a highly variable species native to the Great Basin and Colombia Basin, which can have either yellow or white flowers (there is also a rare orange form).
When attempting to identify some of the yellow-flowered taxa in the group on iNat, I noticed that the differences between the three (R. velutinum, R. quercetorum, and R. lasianthum) were less than clear. Indeed, it should only take one look at the below image which shows the flowers of all 3 side-by-side to realize that distinguishing them is not a trivial task. (they do not look identical of course, but it is hard to say whether the differences are actually substantial or just part of natural phenotypic variation within a single species.)

Lower right side image credit to Steve Matson: https://www.inaturalist.org/observations/109069329
The Jepson and FNA keys both primarily use the traits of:
- hypanthium length,
- hypanthium width, and
- ovary texture
to distinguish between them, but my own personal observations combined with the below two articles made me skeptical that these were sufficient to fully distinguish the three.
https://tchester.org/plants/analysis/ribes/quercetorum.html
https://tchester.org/plants/analysis/ribes/velutinum.html
Indeed, the results of this project indicate that although these three traits have some diagnostic power, they are often insufficient to distinguish between species and must be combined with other factors such as elevation and flowering phenology.
Furthermore, although the initial goal of the study was to investigate the difference between the three yellow-flowered taxa, I also decided to investigate the striking phenotypic diversity of the variable R. velutinum. This species has a wide range of phenotypes in both its flowers and fruit, with the flower color ranging from white to yellow (and rarely orange) and the ovary/fruit texture ranging from densely glandular-hairy to glabrous, and there is even a common form with nonglandular hairs.
So, the overall questions I was aiming to answer with this project were:
- What are the morphological differences between the flowers and/or fruit of the three species, and how can we apply them to identify which species a specimen belongs to?
- Are these morphological differences substantial enough to support the idea that all 3 are distinct species, or do some of the taxa need to be synonymized with the others?
- Conversely, are there yet-undescribed cryptic taxa within the group awaiting formal description?
- Are the traits of flower color and ovary/fruit texture in R. velutinum geographically correlated, and should infraspecific taxa be created or revived in order to better categorize its extreme phenotypic diversity?
METHODS
The project consisted of an analysis of herbarium specimens with the addition of some iNaturalist observations into the data pool (measurements were not taken from any iNat observations, they were used solely for flower color and ovary/fruit texture analysis).
Over 600 specimens loaned from various herbaria were analyzed, with me taking note of the ovary/fruit texture, flowering phenology, flower color, total flower length, hypanthium length, and hypanthium width. The flowering phenology (i.e. budding, flowering, fruiting, in between) was made according to my best judgement looking at the relative size of the ovaries, how open the flowers were, and how desiccated the flowers looked. Flower color was not recorded for any herbarium specimen unless it was listed by the collector due to the tendency of color to fade over time with pressed plants. Hypanthium length was measured as the distance from where the ovary attaches to the sepal tube to where the sepals differentiate themselves from the tube. Total flower length was measured as the distance from where the ovary attaches to the sepal tube to the far tip of the petals. Although total flower length does not seem to have been referenced so far in any floras or literature, it seems to have diagnostic power roughly equivalent to hypanthium length, with the added versatility that total length is easy to measure in fruiting plants whereas hypanthium length is difficult to impossible to measure in fruiting plants. Hypanthium width was measured as the width of the hypanthium at its widest point near where it attaches to the ovary. Below is an annotated image showing example pixel measurements from a flowering specimen of R. velutinum, although all the specimens used in the study were obviously measured with rulers.

Furthermore, floral measurements were only taken from fully open and non-desiccated flowers that were pressed parallel to the sheet. I did not choose to include petal length in the analysis as I find it hard to interpret (do they mean from the outside, end of the hypanthium to the tip of the petals, or to where the petals actually attach), and because it seemed to differ little between individuals or taxa and thus did not seem to be diagnostically useful. I also chose not to utilize traits such as difference in hair type (i.e. "floccose" vs. "wooly") because they are difficult to quantify and not necessarily taxonomically significant. I also did not include vegetative characters (i.e. degree of leaf pubescence, degree of spininess) in the analysis for similar reasons.
RESULTS
Hypanthium length vs. width
As can be seen in the scatter plot below, there is an extremely high degree of overlap between the taxa for these traits. R. quercetorum and R. lasianthum show near complete overlap in their floral dimensions, and although R. velutinum is slightly removed from those two, there is still a somewhat substantial degree of overlap between it and R. quercetorum.

Contrast this to the degree of reliance the Jepson and FNA keys place on this trait and it should be clear why such confusion around these taxa exists.
FNA Key:
50: Hypanthia 4-5 mm: -- Ribes lasianthum
+: Hypanthia 1-3 mm: -- (2)
51: Ovaries usually densely crisped-puberulent, rarely glabrous; sepals 1-2 mm; petals 1.5-2.5 mm: -- Ribes velutinum
+ Ovaries glabrate; sepals 3 mm; petals 1 mm: -- Ribes quercetorum
Jepson Key:
22: Ovary hairs conspicuous, short and long, glandular and not: -- R. velutinum
22': Ovary hairs generally 0 or inconspicuous, generally short, generally nonglandular: -- (23)
23: Hypanthium ± as long as wide – dry interior mountain slopes, 700–2500 m: -- R. velutinum
23': Hypanthium longer than wide: -- (24)
24: Hypanthium > 3 mm; fruit red; subalpine, alpine mountain tops, 2100–3100 m: -- R. lasianthum
24': Hypanthium < 3 mm; fruit black; rocky slopes, oak-covered foothills, chaparral, < 1350 m: -- R. quercetorum
So we can see that the characteristics of the plants that I observed are not consistent with how they are described in the floras. To begin, we can see that many specimens of R. quercetorum and R. lasianthum have hypanthia which approach being roughly as wide as long, while some of R. velutinum are notably longer than wide, which calls into doubt the strength of keybreak 23 in the Jepson. For the FNA key, it lists R. lasianthum hypanthia as being over 4 mm, but my analysis shows that slightly over half of the individuals sampled have hypanthia under 3 mm. There is a similar problem with the Jepson key, wherein it lists R. quercetorum as having hypanthia under 3 mm, which is again strongly contradicted by my data. I have many more criticisms of these keys but those are for later.
If this chart was all one had to go on, they would almost certainly consider R. quercetorum and R. lasianthum to be the same species, with R. velutinum being a doubtful segregate from those two. So, are there other, more reliable ways to separate these species, or do they need to be synonymized?
Elevation, Habitat & Flowering Phenology
One of the reasons why I prefer the Jepson to the FNA key is that it recognizes the differences in elevation and habitat between the 3 entities. One might erroneously conclude from the hypanthium length/width graph that R. quercetorum and R. lasianthum surely must be the same species. But the two occupy completely different ecological habitats and niches -- R. quercetorum is a winter-blooming shrub of temperate coastal regions, and R. lasianthum is a late summer-blooming shrub of strongly seasonal alpine ranges! Consider the following plots of flower/hypanthium length versus elevation or flower phenology:

Looking at these graphs, we can begin to tease out the dynamics of the group a bit. It appears that it consists of two long-flowered taxa clearly separated by at least a few hundred meters of elevation and a few weeks of flowering time, and one short-flowered taxon that bridges the phenology and elevation gap between the two long-flowered ones, but is mostly separable from those two by its shorter flowers.
To be specific, it is obvious that R. quercetorum and R. lasianthum are different species. Although they are morphologically very similar, the two are completely reproductively isolated by both time and space, with at least ~500 meters of elevation and at least 6 weeks of flowering time preventing interbreeding.
However, it is not as simple to distinguish R. velutinum from those two. R. velutinum seems to manifest as a population with notably shorter flowers than the other two taxa that nonetheless has some overlap with them in both flowering phenology and elevation.
Let's begin with the relationship between R. lasianthum and R. velutinum. One thing you may notice about the R. lasianthum data cluster is the vertical lines of data points in R. lasianthum, which were all from my collecting trip in July and do a pretty good job of showing the population-level variance of R. lasianthum. These specimens generally show shorter flowers than the rest of the samples examined, which may have been due to my efforts to sample multiple different plants from the same populations. It is probable to me that the R. lasianthum populations which flower in May also have flowers that can be as short as the ones that I sampled, which would mean that the two are probably less easily distinguished than the graphs might initially show. Still, the two clusters of these species seem more or less distinct, with most of the unusually long-flowered R. velutinum blooming too early in the year to be R. lasianthum, or being located in habitats that are obviously unfit for R. lasianthum.
The relationship between R. quercetorum and R. velutinum is a lot more tricky. On the whole, they still seem to form two distinctive clumps: R. quercetorum being a lower-elevation, early-flowering cluster with a wide range of possible flower sizes that are generally larger than those seen in the other cluster, and with R. velutinum being a higher-elevation, later-flowering cluster that has a more tight distribution of shorter flowers. And although there is still a substantial degree of overlap, the clusters seem distinct enough to warrant the distinction. Furthermore, many of these ambiguous cases can be easily resolved through the use of habitat and geography. However, there are still a few regions where the distinction between these two remains not fully clear, which I will address later, at the end of this post.
Overall, the data supports the recognition of three relatively distinct (species) lineages within this group, but in order to make this distinction we need to reference more traits (such as elevation and phenology) than solely floral dimensions.
Fruit Characters: Color and Floral Remnant Length
Another issue I have with the Jepson key is how it references fruit color as one of the distinguishing traits between R. velutinum and R. lasianthum. The main problem with this is how it ignores that fruit color can vary between individuals of a species, and even between the same individual depending on the maturity of the fruit! Consider the below photo of R. quercetorum, showing a branch that contains fruit with a variety of colors depending on their maturity, starting at green when young to yellow then red and finally black when mature:

Photo Credit gilbertj: https://www.inaturalist.org/observations/70961739
You can also look at the below collage of R. velutinum fruit to see an equal amount of variation in that species:

Photos Credit J. Stauffer (top and bottom left) and R.J. Adams (right), respectively: https://www.inaturalist.org/observations/36866256, https://www.inaturalist.org/observations/36883641, https://www.inaturalist.org/observations/3670653
Unfortunately, there don't seem to be many photos of R. lasianthum fruit, but we can read between the lines here to infer that the fruits are equally diverse. But regardless of the possible fruit colors that R. lasianthum actually has, the mere fact that both other yellow-flowered species in the group can have both red or black fruits means that it is effectively useless as a diagnostic trait.
So not only can R. velutinum have red fruit, which contradicts the Jepson key, but R. lasianthum presumably follows the same pattern as the rest wherein the red fruits are just less mature than the (fully mature) black ones. I suspect that the phenology and habitat of R. lasianthum may have unconsciously led to this kind of bias: given that its fruits mature very late in the year (late Aug-Sep), the high alpine habitats in which it grows may be less accessible by this time, leading to the association of that taxa to its immature red fruits. Given all these issues, it does not seem like fruit color can be used to reliably separate any of the taxa, and much more research needs to be done into how the fruit color changes over time in all 3 species until we can begin to say that it is a useful diagnostic trait.
Finally, it is also still an open question as to whether the length of the dry floral remnant can be used to distinguish between species, as flowers are not always available for identification. One traditional problem in using this trait seems to be confusion between the length of the hypanthium and the total length of the flower. It can be difficult to discern the length of the hypanthium from just a dried floral remnant, and if one measures from the point of attachment to the berry to where the remnant terminates, they will actually just be measuring the total length of the flower. Thus, it seems to reason that total length should be the metric that is used when evaluating floral remnants, as it offers similar diagnostic power to hypanthium length but is easier to measure. However, the data that I collected is unable to comment on whether the total length of the floral remnant is a reliable diagnostic tool.
Ribes lasianthum and Ribes quercetorum: More phenotypically diverse than previously known
During the analysis, I found multiple individuals of both R. quercetorum and R. lasianthum which had glandular-hairy or densely pubescent ovaries. This is in stark contrast to how both are described in the floras and literature, with both the Jepson, FNA and original descriptions of the two explicitly describing the berries/ovaries as "glabrous" (except for the original description of R. lasianthum, which describes them as "hairy", but this seems to be an error as this phenotype is uncommon in my data). Furthermore, as has been discussed before, the flowers of R. lasianthum are significantly more diverse in size and shape than before described, with shapes ranging from short and stout to long and wide. This diversity in ovary texture and flower shape in R. lasianthum is shown in the bellow collage:

I also prepared maps showing the geographic distribution of ovary texture in these two taxa:

We will get back to these later and discuss the implication this has for potential hybrid zones, but it should suffice to say now that the ovary texture in this group is much more diverse than is stated in the literature, which adds further difficulty to distinguishing them. The keys imply that R. quercetorum and R. lasianthum always have glabrous ovaries, which further implies that any specimen with glandular-hairy ovaries must be R. velutinum, but my data indicates that this is plainly not true. Ovary texture can generally be diagnostically useful in that R. velutinum ovaries tend to be more glandular than those in R. quercetorum, but it is not a hard and fast rule and should not be treated as such.
Untangling the many forms of R. velutinum: Flower color and ovary texture
Another main goal of the project was to reevaluate the wide range of phenotypic diversity present in R. velutinum. Numerous infraspecific taxa have been proposed over the years in order to accommodate this diversity, with the focus on these names being on the flower color and ovary texture. The most commonly used names seem to be var. glanduliferum, being applied to the individuals with glandular-hairs on the ovaries/fruits, var. gooddingii, being applied to those with glabrous ovaries and deeply lobed leaves, and var. velutinum going to those which do not fit those above characters. Currently, neither POWO, FNA, or Jepson recognize any infraspecific taxa for R. velutinum, and the below map showing the geographical distribution of flower color and ovary texture for the species might indicate why:

We can clearly see that ovary texture is not particularly strongly geographically correlated. There are some weak patterns (the glandular form is less common in the far northern regions, wherein the glabrous form dominates, whereas the glandular form is much more common on the boundary between the Sierras and the Great Basin), but overall the different forms overlap significantly. This would seem like strong evidence that ovary texture is just a highly variable phenotypic trait, which would back up the practice of not recognizing var. glanduliferum, but I also think that more work at the population level needs to be done to determine whether or not each of these different forms ever coexist at in the same population before a definitive conclusion is reached.
The status of var gooddingii is not as simple. From my survey of the loaned herbarium specimens, it seems that this name has been frequently misapplied to plants which do not fit the description (i.e. applied to all plants with glabrous ovaries or to all plants with white flowers). The initial description of R. gooddingi (https://www.biodiversitylibrary.org/item/100225#page/74/mode/1up) clearly notes that it differs from typical R. velutinum not only in the glabrous ovaries but also (critically) the less pubescent leaves and deeper leaf lobes. Also, although var. gooddingii always has white flowers, this does not mean that all individuals of R. velutinum with white flowers are var. gooddingii. Unfortunately, I did not collect any quantitative data about leaf characters in the study, so it is hard for me to make a definitive answer. However, browsing pics on iNat seems to show that the slightly disjunct populations of R. velutinum known from northeastern Oregon, southeastern Washington, and central Idaho have substantially different leaves than those in the populations southwards (compare these two obs of what would and would not, respectively, qualify as var. gooddingii in my interpretation: https://www.inaturalist.org/observations/154639731, https://www.inaturalist.org/observations/72591131). Thus, I have to tentatively disagree with the choice to not recognize var. gooddingii, with the caveat that this variety seems to strictly only apply to the disjunct populations in NE Oregon, SE Washington, and C Idaho with their nearly glabrous, deeply divided leaves.
In contrast, whereas ovary texture does not appear to be geographically correlated, flower color is the exact opposite. Not only is the white form overwhelmingly more common in the northern part of its range (and vice versa for yellow), but there does not appear to be any overlap between the ranges of the two. Also, it is very interesting that the rare orange-red phenotype is known from right at the contact zone between the two more common colors, where it grows sympatrically with the yellow form.
Of course, this dataset is very incomplete -- there are no collections which mention flower color or iNat observations with flower pictures over much of the species' range near the apparent dividing line in the Great Basin, so efforts should be made to document flower color at points throughout this range so that the exact extent of both forms is known. There also needs to be work at populations near the contact zone to see if both color forms can be found growing in the same population. However, even if it can be said that there is some overlap in the ranges between the two color forms, I find this geographic correlation to be strong enough to consider the white and yellow flowered individuals different taxa, at the subspecific level. Such a change allows for clear dividing lines between which subspecies an individual belongs to and is clearly not arbitrary. Of course, I will not be formally proposing a name for this right now (or in the foreseeable future) but I just want to put the argument out there.
"Problem Areas:" Contact zones and potential hybridization
Although all 3 taxa tend to occupy more or less distinct habitats and geographical areas there are some areas where the ranges of two overlap. This becomes problematic in the case of R. quercetorum and R. velutinum in the foothills of the far southern Sierras, right between the coastal areas that favor the former species and the inland desert areas that favor the latter. It appears that both taxa exist in this location, evidenced by the presence of both extremely long (https://www.inaturalist.org/observations/156841748) and extremely short flowered plants here. However, not only does R. quercetorum seem to bloom somewhat later than usual here (probably to the harsher climate than in the rest of its range), but the aforementioned small degree of overlap in the floral dimensions of the two species mean that it may not always be possible to distinguish between the two in this area. It is also possible to me that the two could hybridize and intergrade here, although much more work needs to be done at the population level to see if this is true. However, I do not think that it is a coincidence that both individuals of R. quercetorum with glandular-hairy ovaries (and half of the individuals with densely pubescent ovaries) that were sampled were collected from near this contact zone, and view it as potential evidence that gene flow exists between the two species at this location.
Finally, there are two iNat observations from Mt. Pinos (https://www.inaturalist.org/observations/169499295, https://www.inaturalist.org/observations/170864167) where I am uncertain of their identity. The high, rocky alpine habitat and late flowering date seems to better match with R. lasianthum, but it is hard to tell exactly how long the flowers are from the pics, and the phenology is in a bit of a grey area to the point where I could imagine these being R. velutinum. It would be interesting to see collections from this locality which get precise floral measurements in order to make a better determination on which species this is.
Final takeaways and a revised key
I think this post is long enough already, so let's wrap it up. Although R. velutinum, R. lasianthum, and R. quercetorum seem to be distinct species, their superficial similarities, combined with many errors in the keys and floras for the three, have led to some confusion about how to distinguish them. Below is a main summation of the problems with this group so far:
- The keys state that the floral dimensions of R. quercetorum and R. lasianthum are distinct enough to be used as a distinguishing factor between the two, but this is not true. In fact, their floral dimensions show almost complete overlap.
- Furthermore, the floral measurements given in both keys are not always correct, with the hypanthia of R. quercetorum having the potential to be much longer than in the keys, and with those of R. lasianthum having the potential to be much shorter.
- Elevation, phenology, and habitat are highly valuable diagnostic tools, especially they when combined with floral dimensions. However, the FNA key references none of these traits, and the Jepson does not reference phenology, to both of their detriments.
- The trait of hypanthium length:width ratio is generally a reliable way to distinguish R. velutinum from the other two species, except in the case of extreme outliers (see Addenda for more information)
- The Jepson key uses fruit color as a distinguishing trait between R. lasianthum and R. velutinum, despite evidence showing this to be inconsistent and confusing.
- R. quercetorum and R. lasianthum are both only described as having glabrous ovaries/fruits in the floras. However, both species can occasionally possess glandular-hairy to pubescent ovaries, with the glandular-hairy form actually being rather common in R. lasianthum.
- Because of this, the first keybreak in the Jepson is erroneous in that it states that all individuals with non-glabrous ovaries are R. velutinum, and a similar error is seen in the second keybreak in the FNA.
- Ovary texture in R. velutinum is not strictly geographically correlated, which corroborates the idea of not recognizing var. glanduliferum.
- In contrast, flower color shows extreme geographic correlation, giving strong support to the notion that a new name for the white-flowered form should be introduced at some subspecific rank.
Given these issues, it is not surprising that many have found the group difficult to interpret. Despite this, the error rate for specimens of these species is surprisingly low. Out of the 628 loaned specimens I examined, only 26 of them were misidentified, and 10 of these misidentifications were for a species that was not even one of the 3 species being studied (most of these were specimens that bore the name of R. velutinum but were actually the completely distinct R. montigenum). Conversely, it is also plausible to me that many institutions still possess specimens of R. lasianthum or R. velutinum that are incorrectly labelled as R. montigenum. Overall, the error rate of identification for this group is only 4.14%, which seems to be lower than average, and may indicate that the species are actually better known than previously thought.
Given the results of my study, I will now propose a revised key that both accurately reflects the characters of the plants as they occur in nature, and tries to eliminate the possibility for confusion between the taxa. Also, although the first keybreak relies heavily on flowering phenology, it is a bit more flexible than it initially seems. Obviously, shrubs such as these do not flower all at once in a single day; the flowers are spread out during a period of many weeks, so visiting the same shrub multiple times could help one get a better idea of which species it slots into. Another more practical method could be to check how whether the plant is just starting to flower or if it is just finishing up by looking for flower buds or immature fruit, because a plant that is just starting to flower in June would be easily identifiable as R. lasianthum, whereas a plant that is just finishing up its flowers at the same time would be R. velutinum.
Finally, before the actual key, just a quick review of each of the species as suggested by tchester:
-
R. lasianthum is a late summer-blooming (late May to August) species of strongly-seasonal alpine ranges.
-
R. quercetorum is a lower-elevation, early-flowering (Dec-Mar, rarely to mid Apr) species of temperate coastal regions, with a wide range of flower sizes that are generally larger than those seen in R. velutinum.
-
R. velutinum is a mid-elevation, later-flowering (Apr-mid June) species of inland regions, with a more tight distribution of flowers shorter than those seen in R. velutinum.
1: Flowering Dec-Apr; < 1700 m: -- (2)
+: Flowering May-Aug; > 1700 m: -- (3)
2 (1): Flowers yellow; flowering from Dec-Mar, rarely to mid Apr; 0-1500 m; hypanthia 2-5 mm long, flowers 4-7 mm long; plants of coastal chaparral communities: -- Ribes quercetorum
+ Flowers yellow or white; flowering from late Mar-Jun; > 700 m; hypanthia 1-3 mm long, flowers 3-5.2 mm long; plants of interior desert slopes: -- Ribes velutinum (in part)
3 (1): Flowering from late May-Aug; > 1900 m; hypanthia 2.8-5 mm long, flowers 5-8 mm long; plants spreading, prostrate shrubs of high rocky alpine to subalpine slopes, mostly in the high Sierras, and often near glacial lakes: -- Ribes lasianthum
+ Flowering from Apr-mid June; < 2900 m; hypanthia 1-3 mm long, flowers 3-5.2 mm long; plant habit and habitat various, but usually of hotter, drier, and lower sites than R. lasianthum, and when high elevation, of mountains in the Great Basin or the eastern slope of the Sierras, but never of the high Sierras themselves: -- Ribes velutinum (in part)
If you have any questions feel free to put them in the comments below and I'll try to address them. Lastly, I'd like to give special thanks to those who helped me complete this project, such as Mitch Power and especially Allison Izaksonas at the Garret Herbarium for their assistance, and notably the late Ann Kelsey for establishing the fund who made the project happen.
ADDENDA:
Multiple people requested to see a graph of Elevation vs. Flowering Date, so I included it in the composite image below. As we would expect, there is a generally some correlation between the two, with higher elevation plants blooming later, likely due to climatic factors. However, if we hone in on individual species in this graph, particularly R. quercetorum, the correlation seems to disappear; R. quercetorum seems to bloom irrespective of elevation.
I was also curious about the potential relationship between latitude and phenology; we might expect that plants which are further north might bloom later in the year than those further south due to the difference in seasonal strength correlated with latitude. However, the graphs generally refute this concept, with little obvious correlation existing between the two factors. Despite this, if we hone in on R. velutinum in this plot, we can see a somewhat interesting trend -- it seems as though the populations which are north of the 40th parallel start blooming later in the year but finish blooming earlier than those south of the same line. This is probably a reflection of how R. velutinum has significantly lower variation in elevation in the more northern parts of its range, so those plants all bloom together, unlike the more central Great Basin populations which have drastic variation in elevation and thus have more dispersed bloom times.
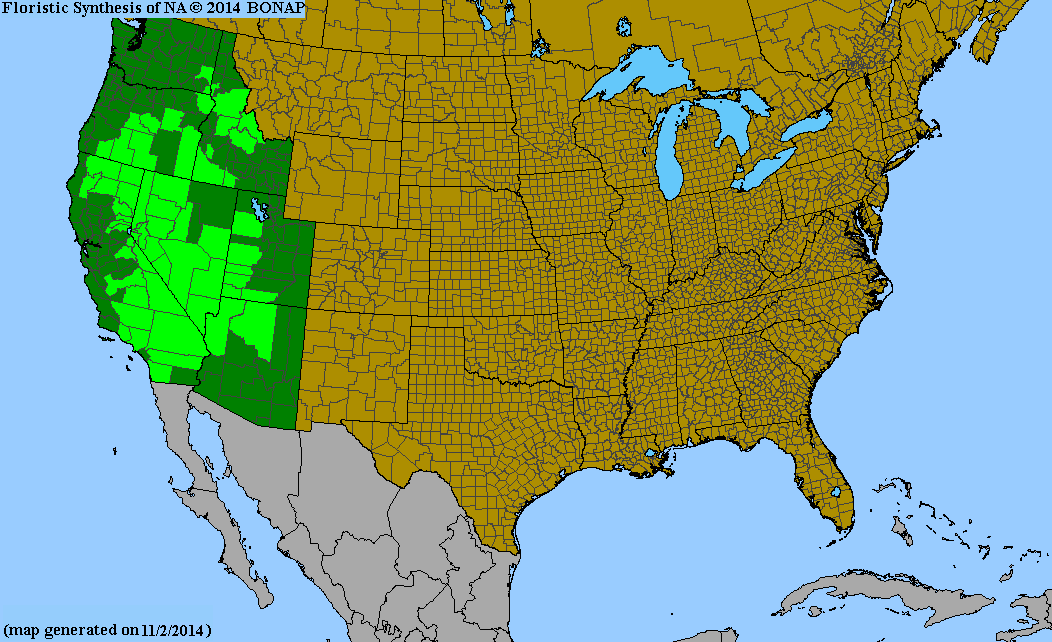
Finally, many people requested a map showing the range of all 3 species as I defined them, which is also included in the below composite. However, note that there were 2 specimens where I could not decide whether to assign them to R. quercetorum and R. lasianthum due to ambiguous elevation, phenology, and a lack of flowers; those specimens are marked with pink dots, as in the legend. Also, this map does not include any areas north of the northern Nevada border, due to it being redundant for comparison purposes (R. velutinum is the only species which grows north of this line).

Finally, there was a request to include box plots for the floral measurement ratios in order to see how reliable this metric is in distinguishing specimens, especially because ratios can be applied to images without scale markers. I went a step further and also included box plots for the objective flower and hypanthium length measurements because they really should have been in the original release. Looking at these plots, they definitely reinforce the concept that all 3 taxa here are separate species because they highlight how much of the measurement overlap between species is due to outliers; accordingly, to help visualize the impact of the outliers on the plots, I decided to place hash marks on the whiskers at the 9th and 91st percentiles, with the whiskers terminating at the min and max.
So we can see from these plots that both the length/width ratios and the objective length measurements are excellent at distinguishing the species in the majority of cases, due to the fact that most of the overlap is beyond the hash marks at the 9th and 91st percentiles. Of course, there is substantial overlap in this category between R. quercetorum and R. lasianthum, but we've already discussed how those two can easily be separated based on elevation and phenology. R. velutinum, meanwhile, shows complete separation from the other species in its ratios, and majority separation in its objective measurements. Overall, because most of the overlap in the ratios in these plots is due to outliers beyond the 9th and 91st percentiles, I will end by concluding that hypanthium length:width ratio is a usually reliable but not infallible way to distinguish R. velutinum from the other two species.







{kind=link}
{kind=link}